ヒストンH4(英: histone H4)は、真核生物の細胞においてクロマチン構造に関与する5つの主要なヒストンタンパク質のうちの1つである。ヒストンH4は主要な球状ドメインと長いN末端テールを持ち、ヌクレオソームのbeads-on-a-string構造の組織化に関与している。ヒストンタンパク質は高度な翻訳後修飾を受ける。共有結合的な修飾には、N末端テールへのアセチル化やメチル化などがある。こうした修飾はヒストンが位置する遺伝子の発現を変化させる可能性がある。ヒストンH4はクロマチンの構造と機能に重要なタンパク質であり、その配列バリアントや多様な修飾状態は遺伝子の動的かつ長期的な調節に関与していると考えられている。
遺伝子
ヒストンH4はさまざまな遺伝子座の複数の遺伝子によってコードされている。
ヒトではHIST1H4A、HIST1H4B、HIST1H4C、HIST1H4D、HIST1H4E、HIST1H4F、HIST1H4G、HIST1H4H、HIST1H4I、HIST1H4J、HIST1H4K、HIST1H4L、HIST2H4A、HIST2H4B、HIST4H4などの遺伝子がヒストンH4をコードしている。
進化
ヒストンタンパク質は、真核生物のタンパク質の中でも最も高度に保存されているタンパク質群である。例えば、エンドウとウシのヒストンH4のアミノ酸配列は102個のアミノ酸のうち2か所しか異ならない。この進化的保存性は、ほぼすべてのアミノ酸がヒストンタンパク質の機能に関与しており、いかなる変化も細胞にとって有害であることを示唆している。ヒストン配列の変化の大部分は致死的であり、致死的でないわずかの変化も遺伝子発現パターンの変化や他の異常を引き起こす。
構造
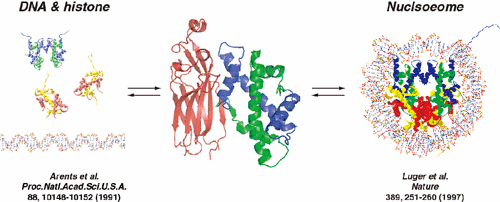
ヒストンH4は102アミノ酸から135アミノ酸からなるタンパク質で、共通してヒストンフォールドと呼ばれる構造モチーフを持つ。ヒストンフォールドは、2つのループで連結された3つのαヘリックスから形成される。ヒストンH3とヒストンH4はH3-H4二量体を形成し、2つのH3-H4二量体が四量体を形成する。この四量体はさらに2つのH2A-H2B二量体を結合し、コンパクトなヒストン八量体コアを形成する。
配列バリアント
ヒストンは最も進化の遅いタンパク質の1つである。ヒストンH4には配列バリアントが存在せず、H4の遺伝子は細胞周期を通じて構成的に発現して同一な配列をコードすると考えられてきた。しかし近年になって、ヒト科特異的バリアントH4Gが同定された。
代替的翻訳
骨形成性成長ペプチド(osteogenic growth peptide、OGP)は、ヒストンH4のmRNAから代替的翻訳によって産生される14アミノ酸からなるペプチドであり、ヒストンH4とC末端の配列ALKRQGRTLYGFGGを共有している。翻訳はヒストンH4のmRNAの85番目のアミノ酸から開始され、19アミノ酸からなるペプチド(preOGP)が合成される。そして、N末端の5アミノ酸が切除されることでOGPに変換される。OGPは、ヒトとラットの血液と再生中の骨髄で確認されている。血清中では、α2-マクログロブリンと他の2種類の未同定のタンパク質に結合した状態で存在している。特異的受容体は同定されていないが、その骨再生機能には一部のシグナル伝達経路が関与していることが明らかにされている。
翻訳後修飾
ヒストンのN末端テールに対するさまざまな共有結合的修飾は、高等真核生物のDNAの機能に必要なさまざまなクロマチン構造の形成を可能にする。メチル化(モノ-、ジ-、トリメチル化)やアセチル化などの修飾がテールに対して行われる。
メチル化
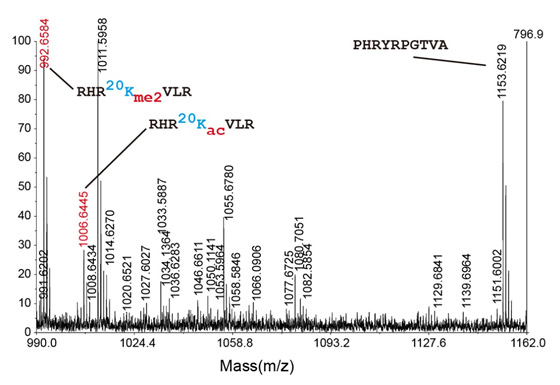
ヒストンのメチル化はアルギニン、リジン、ヒスチジン残基に対して行われる。ヒストンH2A、H3、H4に対するモノ-、ジ-、トリメチル化が発見されている。ヒストンのメチル化は、転写、DNA複製、DNA修復などのDNA損傷応答、ヘテロクロマチン形成、体細胞のリプログラミングなど、さまざまな細胞機能と関係している。これらの生物学的機能の中でも、転写抑制と活性化が最もよく研究されている。PRMT1(ヒストンメチルトランスフェラーゼ)によるH4のR3のメチル化は、in vivoでの広範囲の活性化型クロマチン修飾の確立と維持に必要不可欠なようであることが研究から示されている。また、PRMT1によるヒストンH4のメチル化はその後のN末端テールのアセチル化の許容に十分である。しかしながら、H4のアセチル化はPRMT1によるメチル化を阻害する。
アセチル化
アセチル基の負電荷はDNAのリン酸骨格の電荷と反発してヒストンのDNAに対する結合親和性を低下させるため、ヒストンのアセチル化は凝縮したヘテロクロマチン構造を緩和すると考えられている。この仮説は、いくつかの転写活性化複合体にヒストンアセチルトランスフェラーゼ(HAT)活性が発見されたことで検証された。ヒストンのアセチル化はいくつかの方法でクロマチン構造に影響を与える。第一に、ヒストンのアセチル化はアセチル化ヒストンテールを認識するドメインを持つタンパク質が結合するためのタグとなる。第二に、ヒストンのアセチル化はクロマチンリモデリング因子の機能を防ぐ。第三に、リジン残基の正電荷を中和する。ヒストンH4のK16のアセチル化(H4K16ac)はさまざまな真核生物のクロマチンの構造と機能に特に重要であり、特異的なHATによって触媒される。プロジェリアのモデルマウスにおいて、H4K16の低アセチル化はDNA損傷部位へのDNA修復タンパク質のリクルートの遅れを引き起こすようである。H4K16acは転写活性化とユークロマチンの維持にも関与している。
出典
関連項目
- ヒストンコード
- ヌクレオソーム
- ヒストン
- クロマチン
- クロマチンに関係する他のヒストンタンパク質
- H1
- H2A
- H2B
- H3